por Ronny Nalin
¿Es el Homo naledi nuestro “pariente”, “antepasado”, o parte del “árbol genealógico humano”?

El 10 de septiembre de 2015, un artículo de libre acceso en la revista digital eLife (puede descargarse en http://elifesciences.org/content/4/e09560) presentó a la comunidad científica la adición más reciente de una especie al registro fósil homínido: Homo naledi (Fig. 1).1 Esta presentación incluía la emisión de un documental de casi dos horas de duración producido por PBS y National Geographic titulado “El Amanecer de la Humanidad”, así como la entusiasta divulgación del sensacional descubrimiento por parte de los principales medios de comunicación.
El material fósil descrito en el trabajo es efectivamente extraordinario por varias razones. Ante todo, el número absoluto de restos esqueléticos y dentales recuperado (por encima de los 1500, con probabilidad de aumentar al continuar excavando en el mismo sitio) lo convierte en “la colección más grande de una sola clase de homínidos que haya sido descubierta en África hasta ahora”.2 Un mínimo de 15 individuos está representado en estos restos, abarcando grupos de edades diferentes, desde infantes hasta adultos mayores. Los restos están bastante bien conservados, con alguna rotura y abrasión superficial, y en general están desarticulados excepto en algunos casos en particular (por ej., los huesos de una mano y los de un miembro inferior todavía articulados).
El segundo aspecto que llamó la atención en las noticias es la mezcla de rasgos anatómicos similares a humanos y a australopitecos que exhiben algunas partes recuperadas (por ej., “manos y pies semejantes a humanos” y “pelvis semejantes a australopitecos”).3 Finalmente, el contexto en el que los fósiles están conservados también es sumamente intrigante. Se encontraron los restos en 20 centímetros de fino sedimento que cubrían el piso de una cámara profunda en un sistema de cuevas (llamado Rising Star) cerca de Johannesburgo (Sudáfrica), actualmente accesible sólo a través de pasajes angostos (de hasta 20 centímetros de diámetro) con caídas verticales de más de 10 metros (Fig. 2).
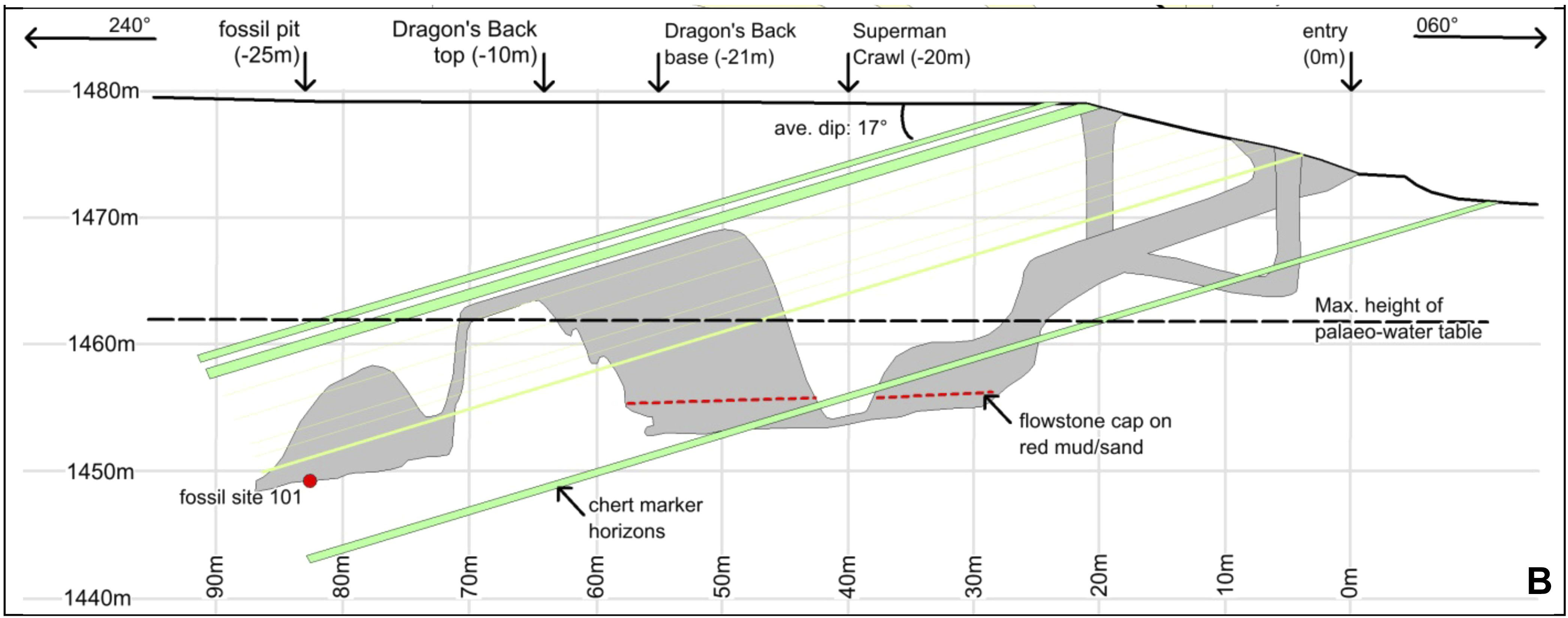
No parece que la cámara cavernaria tuviera otros accesos en el pasado ni que el agua hubiera transportado los restos a la cámara. Además, el conjunto consta exclusivamente de restos de homínidos, sin restos de ningún otro animal mezclados con ellos (con excepción de los huesos de un búho y algunos dientes de roedores). Esta configuración condujo a especular que los cadáveres podrían haber sido dispuestos intencionalmente en este hueco profundo y oscuro, como se debate en un artículo acompañante en eLife (puede descargarse en http://elifesciences.org/content/4/e09561).4
Para aquellos que valoran el conocimiento, la ciencia, y la Palabra de Dios, descubrimientos así evocan siempre una reacción ambivalente. Estamos asombrados por las nuevas evidencias del pasado, pero entristecidos al ser presentados a las masas con el teñido de evolución Darwiniana. Por ejemplo, en sólo unos párrafos, un artículo de CNN sobre el Homo naledi usa expresiones como “nueva especie de antepasado humano”, “una nueva adición a nuestro árbol genealógico”, y “nueva especie de familiar humano”.5 El mensaje implícito en este uso del lenguaje es que los seres humanos no son el resultado de un acto intencional de creación divina, sino meramente una de las muchas secciones del árbol de ascendencia común universal de las formas ancestrales. Esta perspectiva contrasta con el modelo alternativo de un Dios que crea tipos distintos (incluyendo seres humanos) que pudieron modificarse con el paso del tiempo. El lenguaje es una herramienta poderosa que puede usarse para defender interpretaciones específicas de los datos. Sin embargo, por lo general los datos mismos pueden ser compatibles con múltiples hipótesis. El objetivo de este artículo es utilizar el ejemplo del Homo naledi para ilustrar la diferencia entre dato e interpretación, y debatir algunas de las preguntas que un creacionista bíblico podría tener en relación con este nuevo descubrimiento.
Comprendiendo las limitaciones de los datos
El conjunto fósil de Homo naledi es un ejemplo excelente de las limitaciones intrínsecas en la práctica de las ciencias históricas. Podemos trabajar solamente con lo que está conservado y disponible, y desarrollar escenarios plausibles sobre la base de la inferencia más lógica, excluyendo hipótesis sin fundamento. He aquí algunos ejemplos para ilustrar el punto:1) Integridad. En términos de cantidad de restos fósiles y representación de partes del cuerpo diferentes provenientes de un único sitio, el descubrimiento en la cueva Rising Star es muy importante. Sin embargo, incluso a pesar de esta abundancia, el registro todavía es fragmentario. Por ejemplo, hasta ahora se recuperaron restos de al menos 15 individuos diferentes, pero están representados solamente cinco cráneos parciales y algunos otros fragmentos craneales. Algunas partes del cráneo no están conservadas o están documentadas solamente con una muestra. Teniendo en cuenta estas limitaciones, los rasgos que se presentan como diagnósticos para el cráneo de Homo naledi todavía se fundamentan en una base de datos relativamente limitada. El artículo especifica que cuando los especímenes diferentes preservan la misma parte del cráneo, “coinciden estrechamente en todos los detalles morfológicos”6 Sin embargo, también menciona excepciones, donde se interpretan las diferencias entre los cráneos “como relacionadas con el sexo”,6 que son atribuibles a la variabilidad entre individuos masculinos y femeninos. Desafortunadamente, el trabajo no provee una discusión detallada de las diferencias implicadas.
2) Cifras. Es importante comprender la manera en que se obtiene de los fósiles alguna información cuantitativa. Por ejemplo, en la Tabla 1 del artículo7 se indica la capacidad craneal (una medida del tamaño del cerebro) de Homo naledi de alrededor de 513cm3. En la sección de métodos del artículo se ofrece una buena explicación de cómo se obtuvo esta cifra. El proceso supuso fusionar dos juegos de restos craneales en dos modelos 3D diferentes, interpolando áreas donde no había datos disponibles, y luego hacer un promedio entre los dos volúmenes calculados (Fig. 3).
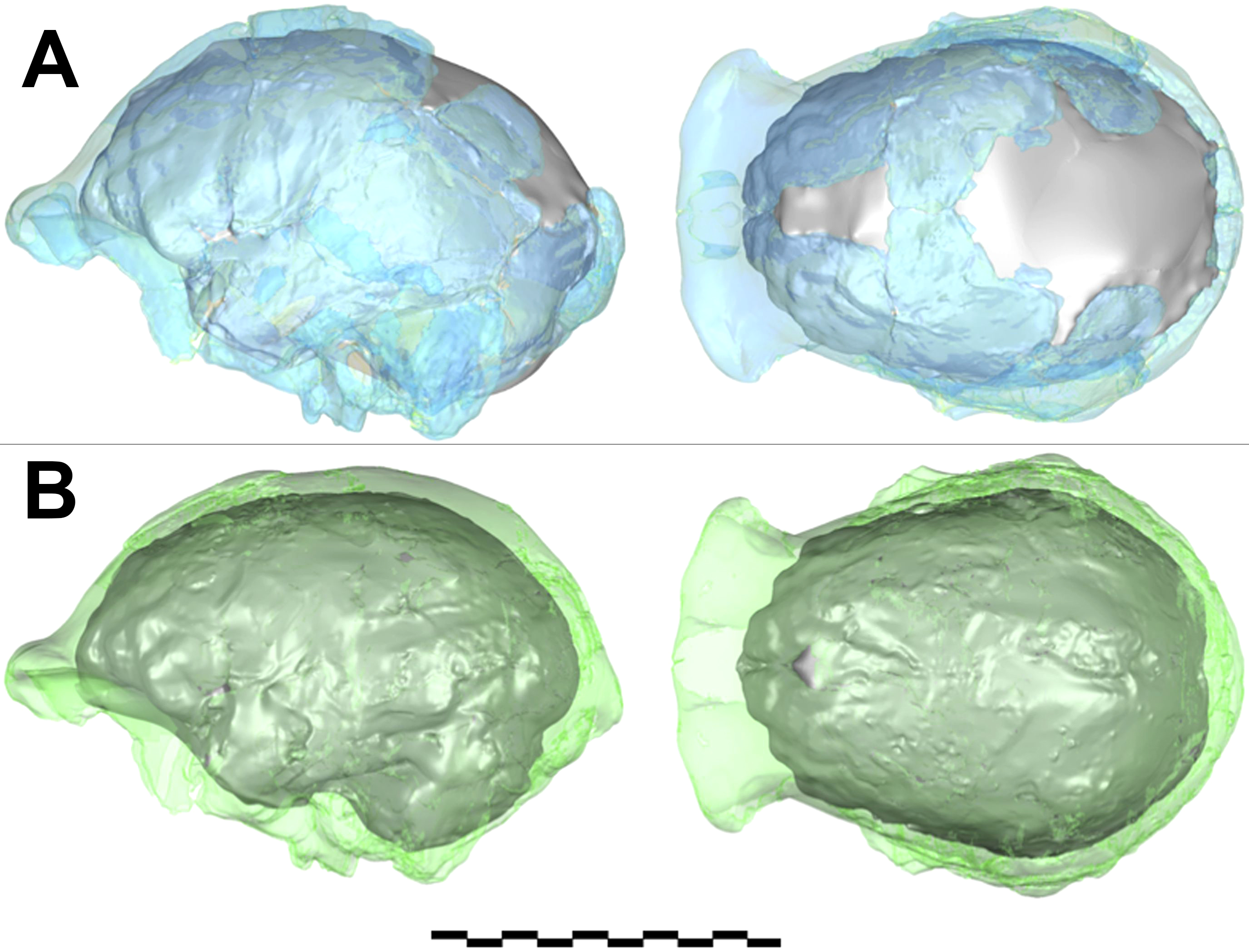
Este proceso de cálculo ofrece una sugerencia bastante buena del tamaño promedio del cerebro que parece representativo de al menos cuatro individuos, pero no es un valor obtenido por la medición directa de un espécimen completo. Otro ejemplo es el cálculo aproximado de la masa corporal. El trabajo estimó la masa corporal de Homo naledi a partir de huesos de ocho individuos, obteniendo un rango entre 39.7 kg y 55.8 kg.8 Se calcularon estos números midiendo la amplitud de los huesos del fémur en ubicaciones estándar y aplicando a las mediciones una ecuación de regresión sobre la base de muestras humanas modernas, donde la masa corporal y la amplitud femoral de individuos humanos pueden establecerse con exactitud y luego correlacionarlas. Éste es el procedimiento estándar, pero está claro que la masa corporal estimada variará si se usan como referencia muestras humanas modernas diferentes y dependiendo del fémur/hueso femoral específico que usaron las mediciones. Por ejemplo, usando una de las mismas mediciones adoptadas en el artículo9 pero con una ecuación de regresión sobre la base de una muestra humana diferente se obtuvo un rango entre 33.09 kg y 40.05 kg,10 que es menor que los valores de masa corporal presentados en el artículo. Además, la suposición de que la correlación entre la amplitud femoral y la masa corporal en los seres humanos modernos es la misma que en homínidos fósiles anatómicamente diferentes obviamente no es verificable, y como máximo probablemente una aproximación.
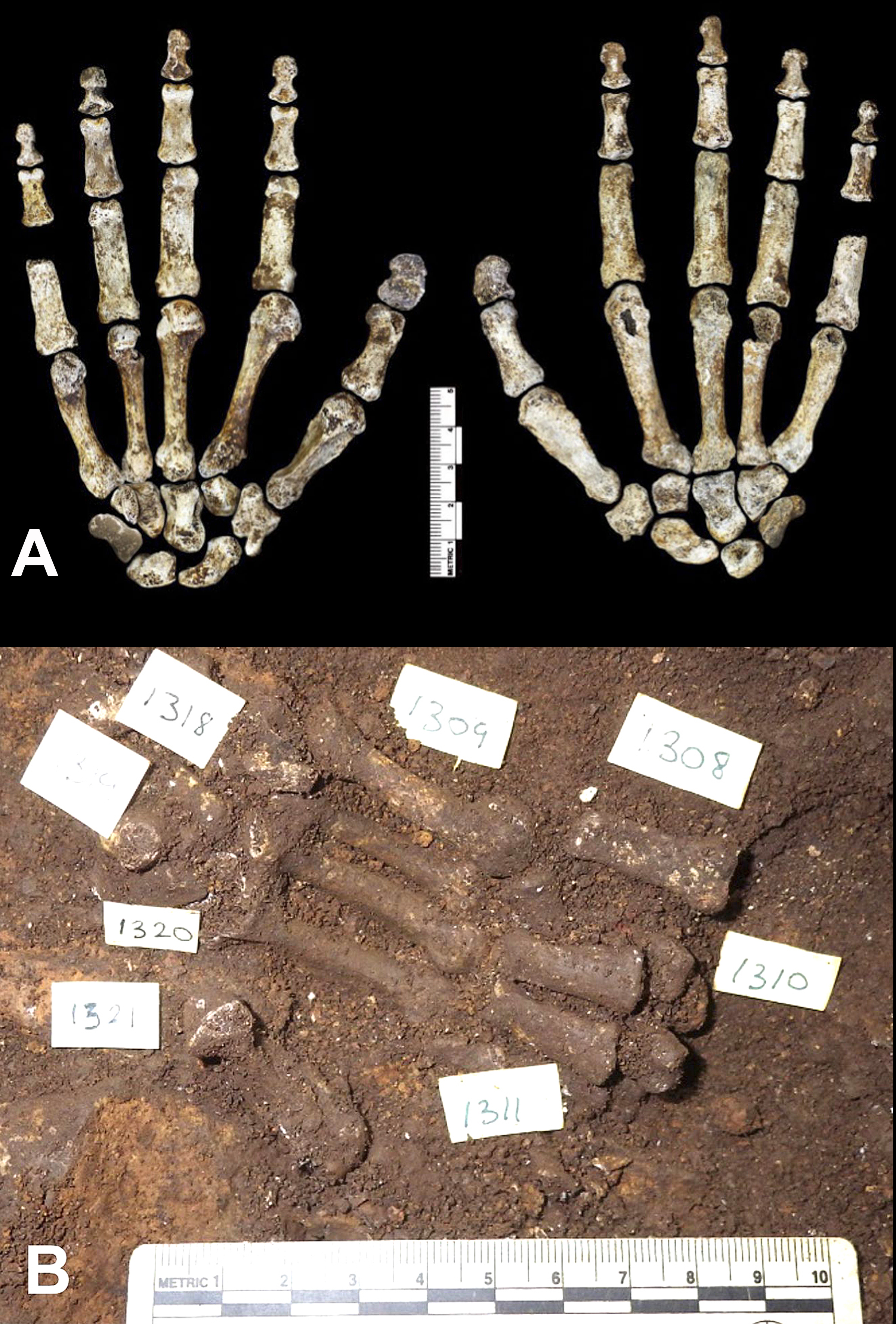
Figura 4 : Los huesos de esta mano son una de las excepciones notables donde se encontraron restos articulados de Homo naledi. A) Vista inferior (izquierda) y superior (derecha) de los huesos de la mano reunidos. Escala en cm. B) Vista de los huesos de la mano articulados durante la excavación. Crédito de la imagen: A) Berger et al. 2015, disponible en http:// elifesciences.org/content/4/e09560; B) Dirks et al. 2015W, disponible en http://elifesciences. org/content/4/e09561#F2
3) Atribución. Dado que la mayoría de los restos estaban desarticulados (ver una excepción en la Fig. 4), es difícil determinar qué huesos pertenecen al mismo individuo. Paradójicamente, si en este conjunto estuviera representado un solo esqueleto, incluso los restos parciales podrían usarse para la medición directa de las verdaderas proporciones del cuerpo. Tener material disociado abundante permite una mejor comprensión del rango de variabilidad morfológico y de tamaño en una población, pero las reconstrucciones de las proporciones del cuerpo pueden ser más complicadas. Otra complicación posible al tratar con restos desarticulados podría resultar en hacer una mezcla de dos tipos distintos de homínidos (por ej. uno más semejante a humanos y otro más semejante a australopitecos) como si pertenecieran a un solo tipo. Los escritores del artículo presentan buenos argumentos para excluir esta posibilidad. Señalan que “en todos los casos donde se repiten varios elementos de área en la muestra, estos son morfológicamente homogéneos, con variación consecuente del tamaño del cuerpo y las diferencias de sexo dentro de una misma población” y que las “configuraciones morfológicas distintivas”, incluyendo “rasgos no encontrados en especies de homínidos ya descritas”, son idénticas en las muestras recuperadas. Su conclusión es que “estas consideraciones indican claramente que este material representa una sola especie, y no un conjunto mezclado”.11 Sin embargo, incluso si este razonamiento es válido, queda la posibilidad teórica de la mezcla de dos tipos, y esto se indicó en los comentarios al descubrimiento realizado por los renombrados paleoantropólogos Jeffrey Schwartz (Universidad de Pittsburgh) y Ian Tattersall (Museo de Historia Natural de Estados Unidos).12
4) Comportamiento. Una demostración final de la diferencia entre los datos y las interpretaciones se observa en la configuración en la que se encontraron los fósiles de Homo naledi. La lejanía de la cámara cavernaria (Fig. 2) y el hecho de que se encontraron solamente restos homínidos llevó a la aseveración de que éste es un ejemplo de disposición intencional de cadáveres. Sin embargo, este escenario es solamente una posibilidad, y no puede establecerse con seguridad. Los escritores reconocen que otros escenarios son posibles: “Tanto los escenarios de una mortalidad masiva o una trampa mortal [...] y la hipótesis de disposición intencional se consideran interpretaciones plausibles y requieren investigación adicional”, aunque los escritores expresan su preferencia hacia la disposición intencional.13 Desafortunadamente, la precaución y el equilibrio expresados en el artículo no se equipararon a los comentarios hechos por los escritores a los medios de comunicación, donde emitieron declaraciones prejuiciadas como éstas: “Acabamos de encontrarnos con otra especie que quizás pensó en su propia mortalidad, y a gran riesgo y esfuerzo dispusieron de sus muertos en una cámara profunda y lejana”;14 “Podemos decir que éste no fue un grupo social que murió por alguna clase de catástrofe en la cueva”;15 “Después de eliminar todas las otras probabilidades, hemos llegado a la conclusión de que Homo naledi estaba utilizando esta recámara en un modo ritual para disponer deliberadamente a sus muertos”.16 Reclamos de comportamiento ritualista, ideas sobre mortalidad propia, y la clara exclusión del escenario de mortalidad masiva de un grupo social no sólo representan una distorsión de las evidencias disponibles, sino también en el último caso están en clara contradicción con las conclusiones del artículo.
La agenda Homo La palabra Homo es altamente relevante, porque es parte del nombre científico de los seres humanos. Si un tipo extinto es puesto dentro del género humano, esto quiere decir que comparte semejanzas más cercanas con los seres humanos que con cualquier otro grupo conocido de organismos, incluyendo los australopitecos. Por lo tanto, la elección de un nombre no es meramente un detalle técnico trivial, sino que expresa un sentido de cómo les gustaría a los descubridores de un fósil que se percibiera el fósil en cuestión. Para los evolucionistas, algo llamado Homo automáticamente ocupa cierto lugar en la supuesta secuencia evolutiva desde los australopitecos hasta los seres humanos. Para los creacionistas, algo llamado Homo encaja automáticamente dentro de las variaciones morfológicas de cierto organismo tipo creado original. Ambos grupos querrían evitar llamar Homo a algo que no lo es.
Los descubridores de los fósiles de la cueva Raising Star tuvieron que abordar dos preguntas cuando escogieron cómo nombrar los restos: 1) ¿representan estos fósiles un nuevo tipo? 2) Si es así, ¿a qué es más similar? Al escoger el nombre Homo naledi, respondieron a la primera pregunta afirmativamente, y sugirieron a los seres humanos como respuesta para la segunda pregunta. Aunque esta elección puede ser legítima, es importante comprender que representa una interpretación específica de los datos, y que otras interpretaciones podrían sugerir conclusiones diferentes a ambas preguntas. Sobre el tema de ser éste un tipo diferente, por ejemplo, el paleoantropólogo Tim White (Universidad de California, Berkeley) considera, de manera provisional, que estos restos podían ser incluidos dentro de la variabilidad de la especie Homo erectus, ya establecida.17
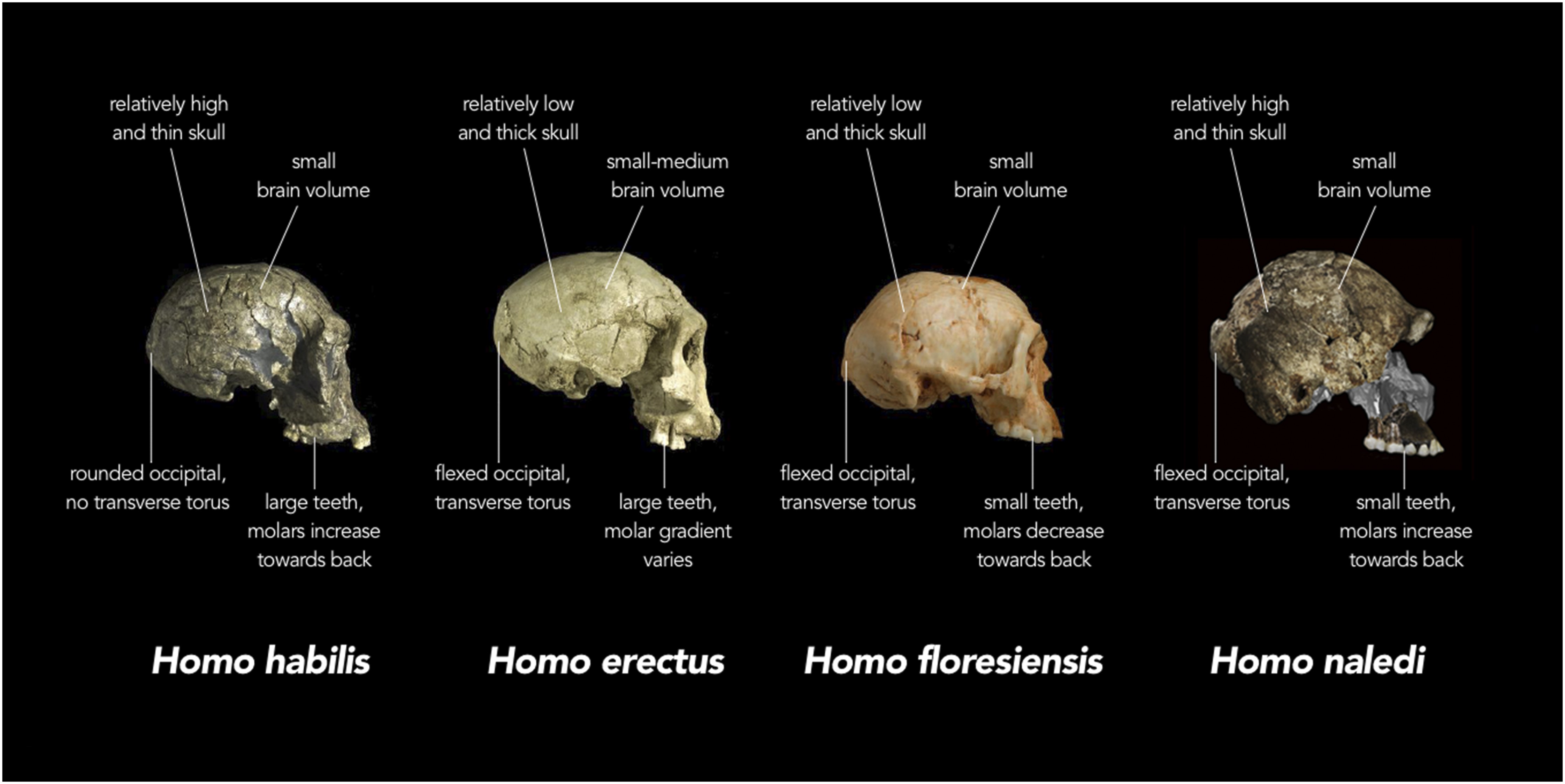
Figura 5 : Cráneos atribuidos a diferentes especies de Homo ilustran el concepto de distribución en mosaico de caracteres. Varias características anatómicas (indicadas con su nombre técnico en la figura) aparecen con diferentes combinaciones en los diferentes especímenes. Crédito de la imagen: http://elifesciences.org/content/4/e10627.
Sobre la elección de la atribución al género Homo, los escritores basan su decisión en los aspectos anatómicos semejantes a los seres humanos descritos de las extremidades inferiores, manos, y dientes, así como por la forma general del cráneo.18 Sin embargo, deliberadamente se pone énfasis sobre estas características mientras que muchos otros rasgos esqueléticos (por ej. la zona pélvica, las región de los hombros, la caja torácica) son más australopitecoides que humanoides. El tamaño aproximado del cerebro, por ejemplo, no sólo es pequeño en valor total cuando se compara con seres humanos modernos, sino que también su proporción relativa a la masa corporal estimada (una medida conocida como cociente de encefalización) es casi la mi- tad de la de los seres humanos modernos y similar a la de los australopitecos.19 En algunas de las presentaciones de los resultados, pareciera que los escritores del artículo querrían deliberadamente enfatizar un vínculo con los seres humanos en vez de con los australopitecos. Por ejemplo, el trabajo dice que “H. naledi tiene un rango de masa corporal similar a las poblaciones humanas de cuerpo pequeño”,2 pero una afirmación igualmente válida habría sido decir que el rango de masa corporal para H. naledi es similar a la masa corporal estimada para los australopitecos grandes.21
Se cree que la glaciación cenozoica se caracterizó por fluctuaciones cíclicas que pueden ser correlacionadas mundialmente (por ej., Zachos et al., 2001). Los geólogos han explicado estas fluctuaciones entre las condiciones glaciales e interglaciales como resultantes de variaciones en los parámetros orbitales de la Tierra (por ej. Maslin et al., 1998; Huybers & Wunsch, 2005; Roe, 2006), que ocurren a escalas de tiempo de 100 ka (excentricidad), 40 ka (oblicuidad) y 20 ka (precesión). Estos rangos de tiempo son problemáticos desde la perspectiva de una cronología breve, y los creacionistas tienen una motivación para buscar procesos alternativos que puedan explicar esta señalización cíclica global dentro de una escala de tiempo mucho más breve.
En conclusión, el estudio cuidadoso de estructuras y depósitos modernos de origen glacial es importante para la interpretación de depósitos anteriores semejantes a los de origen glacial. Al mismo tiempo, los creacionistas abordan las evidencias de glaciación en el registro geológico con atención para explorar hipótesis alternativas compatibles con los modelos de cronología breve de la historia de la Tierra.
1Usamos el término “interglacial” para señalar un período cálido con capas de hielo reducidas entre dos períodos fríos con capas de hielo extensas.
2 ka=mil años
Referencias
1 Berger, L.R., et al. 2015, Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife 4.
2 Íbid., p.3.
3 Íbid., pp.3, 24.
4 Dirks, P.H., et al. 2015, Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa. eLife 4.
5 McKenzie, D. and Wende, H., Homo naledi: New Species of Human Ancestor Discovered in South Africa. CNN, 10 de septiembre de 2015, http://www.cnn.com/2015/09/10/africa/ homo-naledi-human-relative-species/.
6 Berger et al., p. 6.
7 Íbid., p. 11.
8 Íbid., p. 18.
9 Espesor subtrocantérico AP, Tabla 3, p. 18 de Berger et al.
10 Grabowski, M., Hatala, K. G., Jungers, W. L., & Richmond, B. G. 2015, Body mass estimates of hominin fossils and the evolution of human body size. Journal of Human Evolution 85: 75- 93.
11 Berger et al., pp. 4,5.
12 Como aparece en Barras, C., New species of extinct human found in cave may rewrite history. New Scientist, 10 de septiembre de 2015, https://www.newscientist.com/article/ mg22730383-700-new-species-extinct- human-found-in-cave-may-rewrite-history/.
13 Dirks et al., p. 30.
14 Presentado en McKenzie and Wende.
15 Presentado en Johnson, M.A., and Jackson, H., Mind Blown: Is Human Ancestor Discovery the Long-Sought Missing Link? NBC News, 10 de septiembre de 2015, http://www.nbcnews. com/science/science-news/mind-blown- human-ancestor-discovery-long-sought- missing-link-n425406.
16 Presentado en Sample, I., Homo naledi: new species of ancient human discovered, claim scientists, The Guardian, 10 de septiembre de 2015, http://www.theguardian.com/ science/2015/sep/10/new-species-of-ancient- human-discovered-claim-scientists.
17 Presentado en Sample y en Hartley, R., Some bones to pick, Times Live, 18 de septiembre de 2015, http://www.timeslive.co.za/ thetimes/2015/09/18/Some-bones-to-pick.
18 Berger et al., p. 23.
19 Usando los cálculos estimados de masa corporal y tamaño cerebral de Berger et al. y datos y ecuaciones finales de Ruff, C.B., Trinkaus, E. and Holliday, T.W. 1997, Body mass and encephalization in Pleistocene Homo. Nature 387: 173-176, el cociente de encefalización estimado para H. naledi es de cerca de 2.4, mientras que para los seres humanos modernos es de aproximadamente 5.3.
20 Berger et al., p. 18.
21 Por ejemplo, cuando las mismas ecuaciones y tipos de medición usados para calcular la masa corporal en Berger et al., fueron aplicadas a AL 333-131 (un hueso fémur de Australopithecus afarensis) el valor obtenido fue ~ 61 kg, lo que excede todos los estimados aproximados de masa corporal para H. naledi. Usando análisis estadístico multivariado, Grabowski et al. calcularon la masa promedio masculino para A. afarensis como 49.5 kg, que coincide en parte con los cálculos estimados también para H. naledi.
22 Véase, por ejemplo, Schwarz, J.H. and Tattersall, I. 2015, Defining the genus Homo. Science 349(6251):931-932.
23 Berger, L. R., de Ruiter, D. J., Churchill, S. E., Schmid, P., Carlson, K. J., Dirks, P. H. and Kibii, J. M. 2010, Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science 328(5975):195-204.
24 Brown, P., Sutikna, T., Morwood, M. J., Soejono, R. P., Saptomo, E. W. and Due, R. A. 2004, A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431(7012):1055-1061.
25 Lordkipanidze, D., de León, M. S. P., Margvelashvili, A., Rak, Y., Rightmire, G. P., Vekua, A. and Zollikofer, C. P. 2013, A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science 342(6156):326-331.
26 Antón, S. C., Potts, R., and Aiello, L. C. 2014. Evolution of early Homo: An integrated biological perspective. Science. 345(6192):1236828-10.
27 Íbid.
28 Schwarz and Tattersall, p. 932.
29 Antón et al.
30 Berger et al., p. 23.
31 Esta es, por ejemplo, la posición asumida por E. Mitchell en un blog de Answers in Genesis (Respuestas en Génesis) sobre el descubrimiento de H. naledi: Is Homo naledi a New Species of Human Ancestor? 12 de septiembre de 2015, https://answersingenesis. org/human-evolution/homo-naledi-new- species-human-ancestor/
32 La última falsificación que conozco, y que me la señaló un amigo recientemente, quien tenía pocas esperanzas sobre su credibilidad, puede encontrarse en http://worldnewsdailyreport. com/5-meter-tall-human-skeleton- unearthed-in-australia/
Allen, P.A., Etienne, J.L., 2008. Sedimentary challenge to Snowball Earth. Nature Geoscience 1(12):817-825.
Arnaud, E., Eyles, C.H., 2002. Glacial influence on Neoproterozoic sedimentation: the Smalfjord Formation, northern Norway. Sedimentology 49:765-788.
Bennett, M.R., Doyle, P., Mather, A.E., 1996. Dropstones: their origin and significance. Palaeogeography. Palaeoclimatology. Palaeoecology 121:331-339.
Bodiselitsch, B., Koeberl, C., Master, S., Reimold, W.U., 2005. Estimating duration and intensity of Neoproterozoic snowball glaciations from Ir anomalies. Science 308:239-242.
CLIPMAP Project Members, 1981. Geological Society of America Map Series MC-36.
Donovan, S.K., Pickerill, R.K., 1997. Dropstones: their origin and significance: a comment. Palaeogeography. Palaeoclimatology. Palaeoecology 131:175-178.
Eyles, N., Januszczak, N., 2007. Syntectonic subacqueous mass flows of the Neoproterozoic Otavi Group, Namibia: where is the evidence of global glaciation? Basin Research 19:179-198.
Hoffman, P.F., Kaufman, A.J., Halverson, G.P., Schrag, D.P., 1998. A Neoproterozoic snowball Earth. Science 281:1342-1346.
Huybers, P., Wunsch, C., 2005. Obliquity pacing of the late Pleistocene glacial terminations. Nature 434:491-494.
Kilner, B., Mac Niocaill, C., Brasier, M., 2005. Low-latitude glaciation in the Neoproterozoic of Oman. Geology 33(5):413-416.
Lambeck, K., Esat, T.M., Potter, E.-K., 2002. Links between climate and sea levels for the past three million years. Nature 419:199-206.
Maslin, M.A., Li, X.S., Loutre, M.F., Berger, A., 1998. The contribution of orbital forcing to the progressive intensification of northern hemisphere glaciation. Quaternary Science Review 17:411-426.
McCall, G.J.H., 2006. The Vendian (Ediacaran) in the geological record: Enigmas in geology’s prelude to the Cambrian explosion. Earth-Science Review 77:1-229.
Oard, M.J., 1990. An Ice Age Caused by The Genesis Flood. Institute for Creation Research, San Diego. 244 pp.
Oard, M.J., 1997. Ancient Ice Ages or Gigantic Submarine Landslides? Creation Research Society, Chino Valley, Arizona. 130 pp.
Peltier, W.R., 1994. Ice age paleotopography. Science 265:195-201.
Picardi G., Plaut, J.J., Biccari, D., Bombaci, O., Calabrese, D., Cartacci, M., Cicchetti, A., Clifford, S.M., Edenhofer, P., Farrell, W.M., Federico, C., Frigeri, A., Gurnett, D.A., Hagfors, T., Heggy, E., Herique, A.